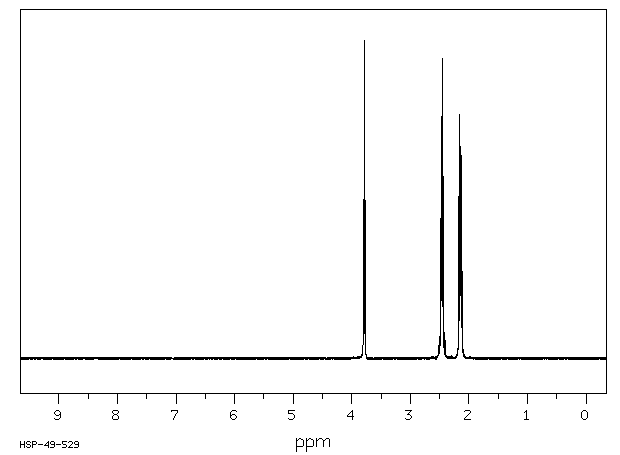
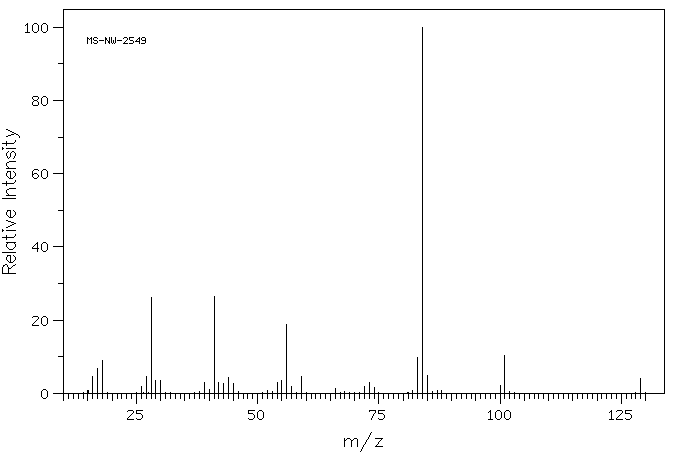
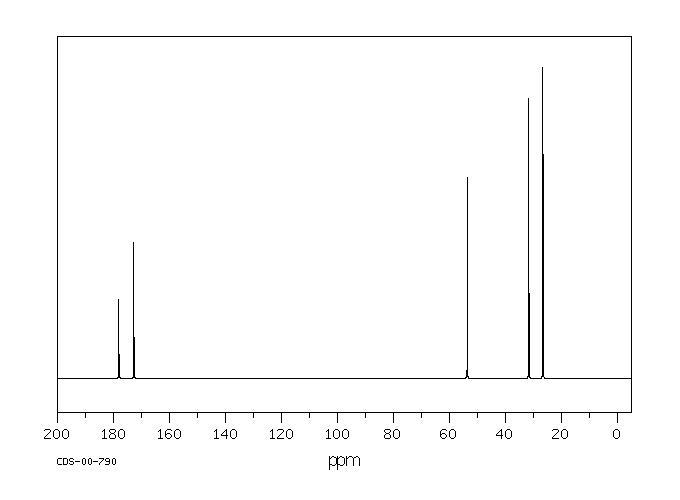
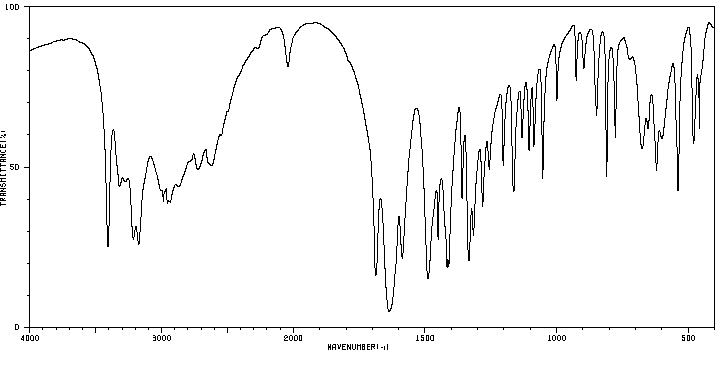
Exogenous L-glutamine likely follows the same metabolic pathways as endogenous L-glutamine which is involved in the formation of glutamate, proteins, nucleotides, and amino acid sugars.
Glutamine plays an important role in nitrogen homeostasis and intestinal substrate supply. It has been suggested that glutamine is a precursor for arginine through an intestinal-renal pathway involving inter-organ transport of citrulline. The importance of intestinal glutamine metabolism for endogenous arginine synthesis in humans, however, has remained unaddressed. The aim of this study was to investigate the intestinal conversion of glutamine to citrulline and the effect of the liver on splanchnic citrulline metabolism in humans. Eight patients undergoing upper gastrointestinal surgery received a primed continuous intravenous infusion of [2-(15)N]glutamine and [ureido-(13)C-(2)H(2)]citrulline. Arterial, portal venous and hepatic venous blood were sampled and portal and hepatic blood flows were measured. Organ specific amino acid uptake (disposal), production and net balance, as well as whole body rates of plasma appearance were calculated according to established methods. The intestines consumed glutamine at a rate that was dependent on glutamine supply. Approximately 13% of glutamine taken up by the intestines was converted to citrulline. Quantitatively glutamine was the only important precursor for intestinal citrulline release. Both glutamine and citrulline were consumed and produced by the liver, but net hepatic flux of both amino acids was not significantly different from zero. Plasma glutamine was the precursor of 80% of plasma citrulline and plasma citrulline in turn was the precursor of 10% of plasma arginine. In conclusion, glutamine is an important precursor for the synthesis of arginine after intestinal conversion to citrulline in humans.
Endogenous glutamine participates in various metabolic activities, including the formation of glutamate, and synthesis of proteins, nucleotides, and amino sugars. Exogenous glutamine is anticipated to undergo similar metabolism.
Supplemental L-glutamine's possible immunomodulatory role may be accounted for in a number of ways. L-glutamine appears to play a major role in protecting the integrity of the gastrointestinal tract and, in particular, the large intestine. During catabolic states, the integrity of the intestinal mucosa may be compromised with consequent increased intestinal permeability and translocation of Gram-negative bacteria from the large intestine into the body. The demand for L-glutamine by the intestine, as well as by cells such as lymphocytes, appears to be much greater than that supplied by skeletal muscle, the major storage tissue for L-glutamine. L-glutamine is the preferred respiratory fuel for enterocytes, colonocytes and lymphocytes. Therefore, supplying supplemental L-glutamine under these conditions may do a number of things. For one, it may reverse the catabolic state by sparing skeletal muscle L-glutamine. It also may inhibit translocation of Gram-negative bacteria from the large intestine. L-glutamine helps maintain secretory IgA, which functions primarily by preventing the attachment of bacteria to mucosal cells. L-glutamine appears to be required to support the proliferation of mitogen-stimulated lymphocytes, as well as the production of interleukin-2 (IL-2) and interferon-gamma (IFN-gamma). It is also required for the maintenance of lymphokine-activated killer cells (LAK). L-glutamine can enhance phagocytosis by neutrophils and monocytes. It can lead to an increased synthesis of glutathione in the intestine, which may also play a role in maintaining the integrity of the intestinal mucosa by ameliorating oxidative stress. The exact mechanism of the possible immunomodulatory action of supplemental L-glutamine, however, remains unclear. It is conceivable that the major effect of L-glutamine occurs at the level of the intestine. Perhaps enteral L-glutamine acts directly on intestine-associated lymphoid tissue and stimulates overall immune function by that mechanism, without passing beyond the splanchnic bed.
In clinical trials of L-glutamine in patients with sickle cell disease, serum aminotransferase elevations were not mentioned, and there were no reports of clinically apparent liver injury. Patients with sickle cell disease frequently have jaundice, largely due to chronic hemolysis which raises serum indirect bilirubin levels. They also can have fluctuating liver test abnormalities due to complications of sickle cell disease, such as gall stone disease (from chronic hemolysis), viral hepatitis and iron overload (from blood transfusions), congestive liver disease (due to pulmonary hypertension), and veno-occlusive crises involving the liver which can be associated with serum aminotransferase elevations and hepatic dysfunction. In preregistration trials of L-glutamine, hepatic events were not reported and serious adverse events were no more common with the active drug than with placebo. L-glutamine is a normal constituent of virtually all tissues and is unlikely to have intrinsic toxicity, even in high doses.
Glutamine supplementation has a potential of worsening hepatic encephalopathy in patients with advanced cirrhosis. Glutamine is metabolized to glutamate and ammonia which can overwhelm the hepatic elimination of ammonia in patients with severe liver dysfunction. Ingestion of 10 to 20 grams of glutamine has been shown to cause elevations of serum ammonia levels and to worsen psychometric measures of hepatic encephalopathy in patients with decompensated cirrhosis. Plasma ammonia levels do not increase with glutamine supplementation in patients with normal hepatic function, and its effects in patients with cirrhosis is not due to hepatic injury. Nevertheless, use of L-glutamine should be avoided in patients with sickle cell disease and advanced cirrhosis.
Likelihood score: E (unlikely cause of acute liver injury with jaundice).
Absorption is efficient and occurs by an active transport mechanism. Tmax is 30 minutes after a single dose. Absorption kinetics following multiple doses has not yet been determined.
Following single dose oral administration of glutamine at 0.1 g/kg to six subjects, mean peak blood glutamine concentration was 1028uM (or 150 mcg/mL) occurring approximately 30 minutes after administration. The pharmacokinetics following multiple oral doses have not been adequately characterized.
Purification and Characterization of Glutamine Synthetase ofPseudomonas taetrolensY-30: An Enzyme Usable for Production of Theanine by Coupling with the Alcoholic Fermentation System of Baker’s Yeast
<i>α-N</i>-Protected dipeptide acids: a simple and efficient synthesis via the easily accessible mixed anhydride method using free amino acids in DMSO and tetrabutylammonium hydroxide
作者:G. Verardo、A. Gorassini
DOI:10.1002/psc.2503
日期:2013.5
to find simple and efficient methods for their synthesis. For this reason, we have investigated the synthesis of α‐N‐protected dipeptide acids by reacting the easily accessible mixedanhydride of α‐N‐protected amino acids with free amino acids under different reaction conditions. The combination of TBA‐OH and DMSO has been found to be the best to overcome the low solubility of amino acids in organic
Anionic chiral cobalt(III) complexes as catalysts of asymmetric synthesis of cyanohydrins
作者:Yu. N. Belokon’、V. I. Maleev、I. L. Mal’fanov、T. F. Savel’eva、N. S. Ikonnikov、A. G. Bulychev、D. L. Usanov、D. A. Kataev、M. North
DOI:10.1007/s11172-006-0338-4
日期:2006.5
Chiral coordinatively saturated cobalt(III) complexes with Schiff bases of enantio-pure amino acids are formed as Λ and Δ-isomers, which are not transformed into each other under normal conditions. These complexes catalyze the formation of enantiomerically enriched cyanohydrins from aldehydes and Me3SiCN under homo-and heterogeneous catalysis.
The present invention provides compounds, compositions thereof, and methods of using the same.
本发明提供了化合物、其组合物以及使用这些化合物的方法。
Microbial enantioselective removal of the N-benzyloxycarbonyl amino protecting group
作者:Michèle Maurs、Francine Acher、Robert Azerad
DOI:10.1016/j.molcatb.2012.03.005
日期:2012.12
Cbz-l-Glu as sole nitrogen source. A lyophilized whole-cell preparation of two Arthrobacter sp. strains grown on Cbz-Glu or Cbz-Gly exhibited a high cleavage activity. The conditions of hydrolysis have been optimized and a quantitative enantioselective deprotection of several Cbz-dl-amino acids was obtained, as well as the deprotection of N-carbamoylester derivatives of several synthetic amino compounds
Potentiometric and Speciation Studies on the Complex Formation Reactions of [Pd(2-methylaminomethyl)-pyridine)(H2O)2]2+ with Some Bio-active Ligands and Displacement Reaction of Coordinated Inosine
作者:Abeer T. Abd El-Karim、Islam R. El-Sherif、Wafaa M. Hosny、Eyad K. Alkhadhairi、Mutlaq S. Aljahdali、Ahmed A. El-Sherif
DOI:10.1007/s10953-017-0621-z
日期:2017.5
of the complexes formed between [Pd(MAMP)(H2O)2]2+ and various biologically relevant ligands containing different functional groups were investigated. The ligands used are amino acids, peptides and DNA constituents. The results show the formation of 1:1 complexes with amino acids and peptides and the corresponding deprotonated amide species. Structural effects of peptides on amidedeprotonation were