
6-氨基-3-甲基嘌呤 | 5142-23-4

-
物化性质
-
计算性质
-
ADMET
-
安全信息
-
SDS
-
制备方法与用途
-
上下游信息
-
文献信息
-
表征谱图
-
同类化合物
-
相关功能分类
-
相关结构分类
物化性质
-
熔点:~300 °C (dec.)(lit.)
-
沸点:240℃
-
密度:1.60
-
闪点:99℃
-
溶解度:可溶于DMSO(高达3mg/ml)或DMF(高达10mg/ml)。
-
稳定性/保质期:
常温常压下稳定,白色至浅黄色或浅粉色的结晶粉末。
计算性质
-
辛醇/水分配系数(LogP):-0.31
-
重原子数:11.0
-
可旋转键数:0.0
-
环数:2.0
-
sp3杂化的碳原子比例:0.166
-
拓扑面积:69.62
-
氢给体数:1.0
-
氢受体数:4.0
安全信息
-
危险品标志:Xn
-
安全说明:S24/25
-
危险类别码:R22
-
WGK Germany:3
-
海关编码:2933990090
-
危险品运输编号:NONH for all modes of transport
-
危险性防范说明:P280,P305+P351+P338
-
危险性描述:H302
-
储存条件:常温、避光、阴凉干燥处,密封保存。
SDS
模块 1. 化学品
1.1 产品标识符
: 3-Methyladenine
产品名称
1.2 鉴别的其他方法
6-Amino-3-methylpurine
1.3 有关的确定了的物质或混合物的用途和建议不适合的用途
仅用于研发。不作为药品、家庭或其它用途。
模块 2. 危险性概述
2.1 GHS-分类
非危险物质或混合物。
2.3 其它危害物 - 无
模块 3. 成分/组成信息
3.1 物 质
: 6-Amino-3-methylpurine
别名
: C6H7N5
分子式
: 149.15 g/mol
分子量
无
模块 4. 急救措施
4.1 必要的急救措施描述
吸入
如果吸入,请将患者移到新鲜空气处。 如呼吸停止,进行人工呼吸。
皮肤接触
用肥皂和大量的水冲洗。
眼睛接触
用水冲洗眼睛作为预防措施。
食入
切勿给失去知觉者通过口喂任何东西。 用水漱口。
4.2 主要症状和影响,急性和迟发效应
据我们所知,此化学,物理和毒性性质尚未经完整的研究。
4.3 及时的医疗处理和所需的特殊处理的说明和指示
无数据资料
模块 5. 消防措施
5.1 灭火介质
灭火方法及灭火剂
用水雾,抗乙醇泡沫,干粉或二氧化碳灭火。
5.2 源于此物质或混合物的特别的危害
碳氧化物, 氮氧化物
5.3 给消防员的建议
如必要的话,戴自给式呼吸器去救火。
5.4 进一步信息
无数据资料
模块 6. 泄露应急处理
6.1 作业人员防护措施、防护装备和应急处置程序
避免粉尘生成。 避免吸入蒸气、烟雾或气体。
6.2 环境保护措施
不要让产品进入下水道。
6.3 泄漏化学品的收容、清除方法及所使用的处置材料
扫掉和铲掉。 放入合适的封闭的容器中待处理。
6.4 参考其他部分
丢弃处理请参阅第13节。
模块 7. 操作处置与储存
7.1 安全操作的注意事项
在有粉尘生成的地方,提供合适的排风设备。
7.2 安全储存的条件,包括任何不兼容性
贮存在阴凉处。 使容器保持密闭,储存在干燥通风处。
7.3 特定用途
无数据资料
模块 8. 接触控制和个体防护
8.1 容许浓度
最高容许浓度
没有已知的国家规定的暴露极限。
8.2 暴露控制
适当的技术控制
常规的工业卫生操作。
个体防护设备
眼/面保护
请使用经官方标准如NIOSH (美国) 或 EN 166(欧盟) 检测与批准的设备防护眼部。
皮肤保护
戴手套取 手套在使用前必须受检查。
请使用合适的方法脱除手套(不要接触手套外部表面),避免任何皮肤部位接触此产品.
使用后请将被污染过的手套根据相关法律法规和有效的实验室规章程序谨慎处理. 请清洗并吹干双手
所选择的保护手套必须符合EU的89/686/EEC规定和从它衍生出来的EN 376标准。
身体保护
根据危险物质的类型,浓度和量,以及特定的工作场所选择身体保护措施。,
防护设备的类型必须根据特定工作场所中的危险物的浓度和数量来选择。
呼吸系统防护
不需要保护呼吸。如需防护粉尘损害,请使用N95型(US)或P1型(EN 143)防尘面具。
呼吸器使用经过测试并通过政府标准如NIOSH(US)或CEN(EU)的呼吸器和零件。
模块 9. 理化特性
9.1 基本的理化特性的信息
a) 外观与性状
形状: 粉末
颜色: 白色
b) 气味
无数据资料
c) 气味阈值
无数据资料
d) pH值
无数据资料
e) 熔点/凝固点
熔点/凝固点: 300 °C - lit.
f) 沸点、初沸点和沸程
无数据资料
g) 闪点
无数据资料
h) 蒸发速率
无数据资料
i) 易燃性(固体,气体)
无数据资料
j) 高的/低的燃烧性或爆炸性限度 无数据资料
k) 蒸气压
无数据资料
l) 蒸汽密度
无数据资料
m) 密度/相对密度
无数据资料
n) 水溶性
无数据资料
o) n-辛醇/水分配系数
无数据资料
p) 自燃温度
无数据资料
q) 分解温度
无数据资料
r) 粘度
无数据资料
模块 10. 稳定性和反应活性
10.1 反应性
无数据资料
10.2 稳定性
无数据资料
10.3 危险反应
无数据资料
10.4 应避免的条件
无数据资料
10.5 不相容的物质
无数据资料
10.6 危险的分解产物
其它分解产物 - 无数据资料
模块 11. 毒理学资料
11.1 毒理学影响的信息
急性毒性
无数据资料
皮肤刺激或腐蚀
无数据资料
眼睛刺激或腐蚀
无数据资料
呼吸道或皮肤过敏
无数据资料
生殖细胞致突变性
无数据资料
致癌性
IARC:
此产品中没有大于或等于 0。1%含量的组分被 IARC鉴别为可能的或肯定的人类致癌物。
生殖毒性
无数据资料
特异性靶器官系统毒性(一次接触)
无数据资料
特异性靶器官系统毒性(反复接触)
无数据资料
吸入危险
无数据资料
潜在的健康影响
吸入 吸入可能有害。 可能引起呼吸道刺激。
摄入 如服入是有害的。
皮肤 通过皮肤吸收可能有害。 可能引起皮肤刺激。
眼睛 可能引起眼睛刺激。
接触后的征兆和症状
据我们所知,此化学,物理和毒性性质尚未经完整的研究。
附加说明
化学物质毒性作用登记: AU6520000
模块 12. 生态学资料
12.1 生态毒性
无数据资料
12.2 持久性和降解性
无数据资料
12.3 潜在的生物累积性
无数据资料
12.4 土壤中的迁移性
无数据资料
12.5 PBT 和 vPvB的结果评价
无数据资料
12.6 其它不良影响
无数据资料
模块 13. 废弃处置
13.1 废物处理方法
产品
将剩余的和不可回收的溶液交给有许可证的公司处理。
受污染的容器和包装
按未用产品处置。
模块 14. 运输信息
14.1 联合国危险货物编号
欧洲陆运危规: - 国际海运危规: - 国际空运危规: -
14.2 联合国运输名称
欧洲陆运危规: 非危险货物
国际海运危规: 非危险货物
国际空运危规: 非危险货物
14.3 运输危险类别
欧洲陆运危规: - 国际海运危规: - 国际空运危规: -
14.4 包裹组
欧洲陆运危规: - 国际海运危规: - 国际空运危规: -
14.5 环境危险
欧洲陆运危规: 否 国际海运危规 国际空运危规: 否
海洋污染物(是/否): 否
14.6 对使用者的特别提醒
无数据资料
模块 15 - 法规信息
N/A
模块16 - 其他信息
N/A
制备方法与用途
3-Methyladenine (3-MA, NSC 66389) 是一种选择性PI3K抑制剂,作用于Vps34和PI3Kγ,在HeLa细胞中IC50分别为25 μM和60 μM。它永久性地抑制I型PI3K,但对III型PI3K的抑制是短暂的,并会阻断自噬体的形成。3-MA可成功应用于抑制线粒体自噬。现配现用(加热助溶)。
靶点| Target | Value |
|---|---|
| Vps34 (HeLa cells) | 25 μM |
| PI3Kγ (HeLa cells) | 60 μM |
3-MA对Vps34的轻微偏向性可能是由于Vps34中某个特异性的疏水环形结构可以环绕在3-MA的3-甲基基团外面造成的。据报道,对于正常培养和饥饿处理的癌细胞,3-MA都能引起细胞死亡。此外,3-MA还可以不通过抑制自噬来抑制细胞迁移和入侵,这表明3-MA除了抑制自噬之外还有其他生物功能。3-MA能够造成半胱天冬酶依赖的细胞死亡,这一功能与其对自噬的抑制无关。用5 mM 3-MA处理葡萄糖饥饿Hela细胞可降低gfp-lc3阳性细胞比例至23%。3-MA处理12到48小时内LC3-I 的水平上升而LC3-II 水平下降,且LC3-I转为LC3-II的过程被3-MA抑制了。2.5 mM 或5 mM 3-MA 处理HeLa 细胞一天并不影响细胞存活率, 而10 mM 3-MA 处理一天会造成细胞存活率下降25.0% 。2.5、5或10 mM 3-MA 处理细胞两天分别使细胞存活率下降 11.5%、38.0% 和79.4%。3-MA降低细胞存活率的作用具有时间和剂量依赖特性。此外,3-MA 明显缩短nocodazole诱导的前中期阻断时间。3-MA通过抑制自噬来抑制SU11274诱导的细胞死亡。在野生型MEF细胞中延长 3-MA处理时间 (最多 9 小时) 明显造成 LC3 I 到 II 转换,且会增加GFP-LC3的聚集而wortMAnnin不具有此功能。3-MA介导的LC3转换和游离 GFP 释放是ATG7依赖的。3-MA处理会导致 p62蛋白水平明显升高。即使在Atg5−/− 的MEF细胞中3-MA也会使p62 水平升高,就像在DOX介导的ATG5缺失的细胞中一样。3-MA以不同方式抑制 I 型和 III型 PI3K。与野生型细胞相比,在Tsc2−/−细胞中3-MA诱导的 LC3 I 到 LC3 II 转换过程大幅下降,且会破坏mTOR复合物1的拮抗自噬功能。
体内研究3-Methyladenine (3-MA)可以通过对磷酸肌醇3磷酸激酶(PI3K)的作用来阻断自噬,而PI3K的活性对于自噬体形成早期膜池的成核和组装是必须的。与SAH处理组相比,3-MA并不会改变出血的程度。然而,与SAH +对照成分组相比经过3-MA预处理后会明显加重神经病学症状。同时,3-MA处理还会减少自噬的发生。相反地,在SAH + 3-MA组里断裂的半胱天冬酶表达量明显上调,与此一致的是与SAH + 对照成分组相比,SAH + 3-MA组中右脑皮层里原位末端标记阳性细胞数量明显增多。
化学性质白色至浅黄色或浅粉色结晶粉末
用途上下游信息
-
上游原料
中文名称 英文名称 CAS号 化学式 分子量 —— 3-Methyl-7-oxy-3H-purin-6-ylamine 155854-89-0 C6H7N5O 165.154 —— 3-benzyladenine 7280-81-1 C12H11N5 225.253 -
下游产品
中文名称 英文名称 CAS号 化学式 分子量 —— 6-Amino-3-methyl-3H-purin-8-ol 185201-03-0 C6H7N5O 165.154 —— 3-Methyl-7-oxy-3H-purin-6-ylamine 155854-89-0 C6H7N5O 165.154 —— 8-Methoxy-3-methyl-3H-purin-6-ylamine 185201-06-3 C7H9N5O 179.181 —— 8-Ethoxy-3-methyl-3H-purin-6-ylamine 185201-07-4 C8H11N5O 193.208 —— 8-Benzyloxy-3-methyl-3H-purin-6-ylamine 185201-08-5 C13H13N5O 255.279
反应信息
-
作为反应物:描述:6-氨基-3-甲基嘌呤 在 sodium hydroxide 、 溶剂黄146 、 间氯过氧苯甲酸 、 硫酸二甲酯 作用下, 以 甲醇 为溶剂, 反应 32.0h, 生成 6-Amino-3-methyl-3H-purin-8-ol参考文献:名称:Purines. LXXIII. Syntheses of 8-Alkoxy- and 8-Hydroxy-3-alkyladenines from 3-Alkyladenine 7-Oxides through 7-Alkoxy-3-alkyladenines.摘要:7- 烷氧基-3-烷基腺嘌呤高氯酸盐(9)由 3-烷基腺嘌呤(4)通过 N-氧化,然后在 N,N-二甲基乙酰胺中用烷基卤化物进行烷基化制备而成。由此得到的 7-甲氧基衍生物 9d、g、j 在沸腾的 0.1 N 氢氧化钠水溶液中分别以 74%、72% 和 39% 的产率得到 3-甲基-8-羟基腺嘌呤(7a)、3-乙基-8-羟基腺嘌呤(7b)和 3-苄基-8-羟基腺嘌呤(7c)。室温下,用甲醇中的甲醇钠处理 9d、g、j,可得到 3-烷基-8-甲氧基腺嘌呤(10m、p、q),收率为 91%-98%。用乙醇中的乙醇钠对 9d 进行类似处理,可得到 8-乙氧基-3-甲基腺嘌呤(10n),收率为 89%。在甲醇存在下,用 0.1 N 氢氧化钠水溶液处理 9d、j,分别以 77% 和 84% 的收率制备出化合物 10m、q。这种方法适用于合成 8-苄氧基化合物 10o:用氢氧化钠水溶液和苄醇的混合物处理 7-苄氧基-3-甲基腺嘌呤高氯酸盐(9f),产率为 60%。化合物 7 可由 9 至 10 交替制备。例如,用甲醇钠处理 9j,然后用沸腾的 1 N 盐酸水解得到 10q,总产率为 84%。另一方面,3-甲基腺嘌呤 7-氧化物(8a)与硫酸二甲酯在 0.1 N 氢氧化钠水溶液中发生甲基化反应,在没有或有添加甲醇的情况下,得到 N6,3-二甲基腺嘌呤 7-氧化物(14),收率为 13% 或 14%,同时得到 7a(收率为 4%)或 10m(收率为 11%)。DOI:10.1248/cpb.44.2025
-
作为产物:描述:3-benzyladenine 在 镍 sodium hydroxide 、 硫酸 、 氢气 、 magnesium monoperoxyphthalate hexahydrate 作用下, 以 甲醇 、 水 、 甲苯 为溶剂, 25.0~40.0 ℃ 、101.33 kPa 条件下, 反应 30.0h, 生成 6-氨基-3-甲基嘌呤参考文献:名称:Fujii, Tozo; Ogawa, Kazuo; Saito, Tohru, Heterocycles, 1994, vol. 38, # 3, p. 477 - 480摘要:DOI:
文献信息
-
[EN] AMINE-LINKED C3-GLUTARIMIDE DEGRONIMERS FOR TARGET PROTEIN DEGRADATION<br/>[FR] DÉGRONIMÈRES DE C3-GLUTARIMIDE LIÉS À UNE AMINE POUR LA DÉGRADATION DE PROTÉINES CIBLES申请人:C4 THERAPEUTICS INC公开号:WO2017197051A1公开(公告)日:2017-11-16This invention provides amine-linked C3-glutarimide Degronimers and Degrons for therapeutic applications as described further herein, and methods of use and compositions thereof as well as methods for their preparation.这项发明提供了胺连接的C3-戊二酰亚胺Degronimers和Degrons,用于治疗应用,如本文进一步描述的,以及它们的使用方法、组合物以及它们的制备方法。
-
[EN] ASH1L INHIBITORS AND METHODS OF TREATMENT THEREWITH<br/>[FR] INHIBITEURS DE ASH1L ET MÉTHODES DE TRAITEMENT AU MOYEN DE CEUX-CI申请人:UNIV MICHIGAN REGENTS公开号:WO2017197240A1公开(公告)日:2017-11-16Provided herein are small molecule inhibitors of ASH1L activity and small molecules that facilitate ASH1L degradation and methods of use thereof for the treatment of disease, including acute leukemia, solid cancers and other diseases dependent on activity of ASH1L.本文提供了ASH1L活性的小分子抑制剂,促进ASH1L降解的小分子以及它们的使用方法,用于治疗疾病,包括急性白血病、实体肿瘤和其他依赖于ASH1L活性的疾病。
-
CONJUGATES OF CEREBLON BINDING COMPOUNDS AND G12C MUTANT KRAS, HRAS OR NRAS PROTEIN MODULATING COMPOUNDS AND METHODS OF USE THEREOF申请人:Araxes Pharma LLC公开号:US20180015087A1公开(公告)日:2018-01-18Conjugates of a cereblon-binding compound and compounds having modulatory activity against G12C mutant KRAS, HRAS or NRAS G12C proteins are provided. Methods associated with preparation and use of such conjugates, pharmaceutical compositions comprising such conjugates and methods to modulate the activity of G12C mutant KRAS, HRAS or NRAS G12C proteins for treatment of disorders, such as cancer, are also provided.提供了与谷氨酰脑结合化合物和具有调节活性对抗G12C突变KRAS、HRAS或NRAS G12C蛋白的化合物的共轭物。还提供了与制备和使用这种共轭物相关的方法,包括含有这种共轭物的药物组合物以及调节G12C突变KRAS、HRAS或NRAS G12C蛋白活性的方法,用于治疗癌症等疾病。
-
[EN] METHODS AND COMPOSITIONS FOR INHIBITION OF RAS<br/>[FR] MÉTHODES ET COMPOSITIONS PERMETTANT L'INHIBITION DE LA RAS申请人:ARAXES PHARMA LLC公开号:WO2016049568A1公开(公告)日:2016-03-31Inhibitors of Ras protein, methods to modulate the activity of Ras protein, and methods of treatment of disorders mediated by Ras protein are provided. A method for regulating activity of a K-Ras, H-Ras or N-Ras mutant protein with a compound is described. Disorders that can be treated include cancer, such as hematological cancer, pancreatic cancer, MYH associated polyposis, colorectal cancer, or lung cancer.提供了Ras蛋白的抑制剂,调节Ras蛋白活性的方法,以及通过Ras蛋白介导的疾病治疗方法。描述了一种使用化合物调节K-Ras、H-Ras或N-Ras突变蛋白活性的方法。可治疗的疾病包括癌症,如血液癌症、胰腺癌、MYH相关性息肉症、结直肠癌或肺癌。
-
[EN] INHIBITORS OF KRAS G12C<br/>[FR] INHIBITEURS DE K-RAS G12C申请人:ARAXES PHARMA LLC公开号:WO2015054572A1公开(公告)日:2015-04-16Compounds having activity as inhibitors of G12C mutant KRAS protein are provided. The compounds have the following structure (I): or a pharmaceutically acceptable salt, tautomer, prodrug or stereoisomer thereof, wherein R1, R2a, R3a, R3b, R4a, R4b, G1, G2, L1, L2, m1, m2, A, B, W, X, Y, Z and E are as defined herein. Methods associated with preparation and use of such compounds, pharmaceutical compositions comprising such compounds and methods to modulate the activity of G12C mutant KRAS protein for treatment of disorders, such as cancer, are also provided.
表征谱图
-
氢谱1HNMR
-
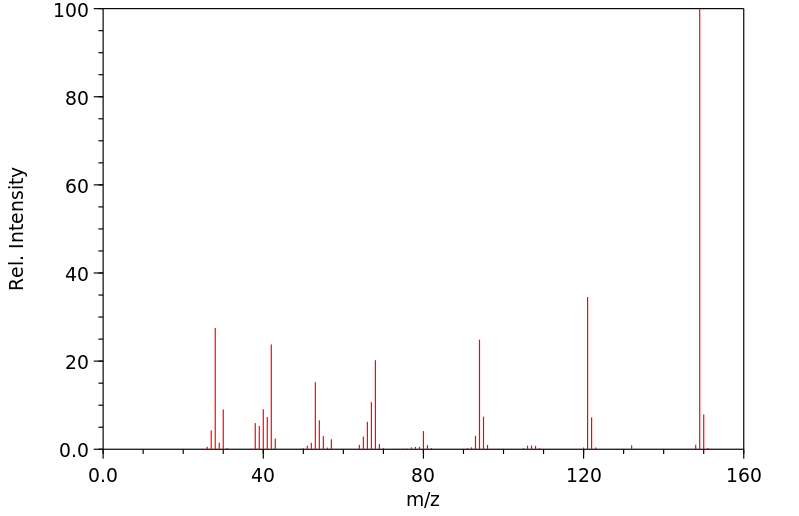
质谱MS
-
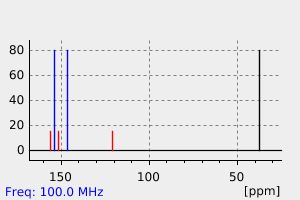
碳谱13CNMR
-
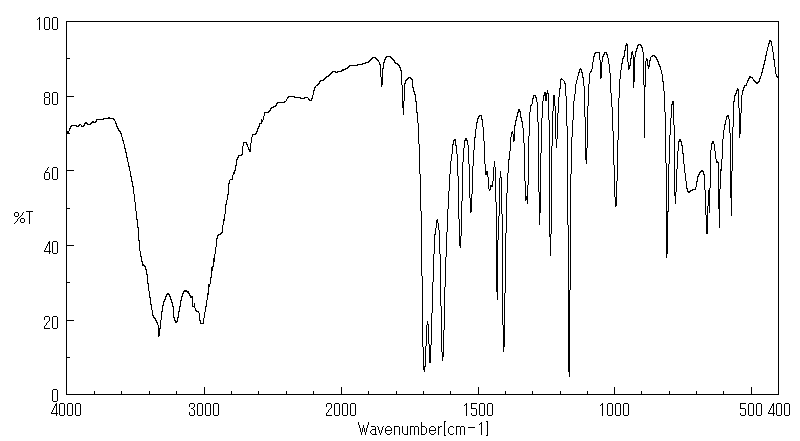
红外IR
-
拉曼Raman
-
峰位数据
-
峰位匹配
-
表征信息
同类化合物
相关功能分类





公众号
